Paralela a la formación del embrión a partir de la gástrula, se desarrollan los anejos embrionarios, unas envolturas de protección llamadas
membranas extraembrionarias.
Tienen especial significado en reptiles, aves y mamíferos. Se trata, por tanto, de un proceso de adaptación al ambiente terrestre.
Estos animales evolucionaron mediante huevos con cubiertas como la cáscara, fárfaras y albúmina, que podían ser dejados en tierra, y con adaptaciones especiales dentro de los huevos que permitían la supervivencia del embrión en el medio terrestre.
Un grupo de células rodea al embrión después de la gastrulación y forma
4 hojas extraembrionarias. Todos los reptiles, aves y los mamíferos actuales poseen estas membranas.
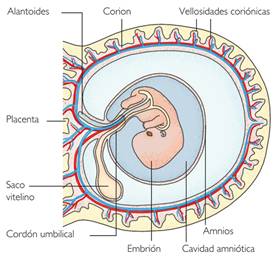
Estas membranas extraembrionarias se denominan:
corion, amnios, alantoides y saco vitelino.
Corion y amnios proceden del ectodermo y mesodermo. Como reptiles, aves y mamíferos tienen amnios, se denominan amniotas, para diferenciarlos de otros vertebrados como peces y anfibios, anamniotas, que carecen de amnios.
- La membrana extraembrionaria más externa es el corion. Evita la evaporación excesiva de agua a través de la cáscara.
- Una segunda membrana, el amnios, rodea al embrión por todas partes menos por la ventral. Delimita la cavidad amniótica, llena del líquido amniótico, que baña al embrión y representa el sustituto del medio acuoso en el que se desarrollaban las larvas de otros animales menos evolucionados y de hábitos más ligados al medio acuático. Además, este líquido amniótico protege al embrión de golpes y otros accidentes.
- Inmediatamente debajo del corion se extiende el alantoides, que es la estructura respiratoria del embrión. El intercambio gaseoso se produce entre numerosos vasos sanguíneos del alantoides y el aire exterior. También sirve para acumular los desechos metabólicos hasta el momento de eclosión del huevo.
- La membrana de la parte ventral es el saco vitelino, que va disminuyendo de tamaño a medida que es consumido por el crecimiento del embrión.
Alantoides y saco vitelino proceden de evaginaciones del endodermo con el mesodermo.